Please refer to Sexual Reproduction in Flowering Plants Class 12 Biology notes and questions with solutions below. These revision notes and important examination questions have been prepared based on the latest Science books for Class 12. You can go through the questions and solutions below which will help you to get better marks in your examinations. We have provided the latest Class 12 Biology Notes and Questions for all chapters in your NCERT Class 12 Biology Book.
Class 12 Biology Sexual Reproduction in Flowering Plants Notes and Questions
All flowering plants (angiosperms) show sexual reproduction. Flowers are the sites of sexual reproduction.
PRE-FERTILISATION: STRUCTURES & EVENTS
- Several hormonal and structural changes result in differentiation & development of the floral primordium.
- Inflorescences bear the floral buds and then the flowers
STRUCTURE OF A FLOWER

A typical flower has 2 parts: Androecium & Gynoecium.
Androecium (male reproductive part)
It consists of a whorl of stamens. Their number and length are variable in different species.
A stamen has 2 parts:
a. Filament: Long and slender stalk. Its proximal end is attached to the thalamus or the petal of the flower.
b. Anther: Terminal and typically bilobed. Each lobe has 2 thecae (dithecous). Often a longitudinal groove runs lengthwise separating the theca.
Transverse section of anther:
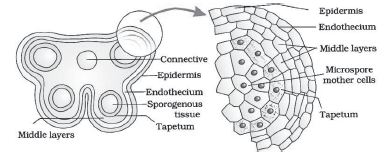
- The anther is a tetragonal structure consisting of four
microsporangia located at the corners (2 in each lobe). - The microsporangia develop to pollen sacs. They extend longitudinally all through the length of an anther and are packed with pollen grains.
Structure of a microsporangium:
- A typical microsporangium is near circular in outline.
- It is surrounded by 4 wall layers: epidermis, endothecium, middle layers & tapetum (innermost layer).
- The outer 3 layers give protection and help in dehiscence of anther to release the pollen.
- The tapetum nourishes the developing pollen grains. Cells of the tapetum contain dense cytoplasm and generally have more than one nucleus.
- In young anther, each microsporangium has sporogenous tissue at centre. It consists of compactly arranged homogenous diploid cells (sporogenous cells).
Microsporogenesis:
- As the anther develops, each sporogenous cell (microspore mother cell or pollen mother cell) undergoes meiotic divisions to form microspore tetrads (microspores arranged in a cluster of four cells).
- Formation of microspores from pollen mother cell (PMC) through meiosis is called microsporogenesis.
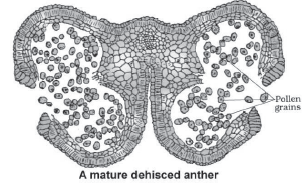
- As the anthers mature and dehydrate, the microspores dissociate from each other and develop into pollen grains.
- Each microsporangium contains thousands of pollen grains.
They are released with the dehiscence of anther.
Pollen grain (male gametophyte):
Generally spherical. 25-50 μm in diameter. Cytoplasm is surrounded by a plasma membrane.
A pollen grain has a two-layered wall: exine and intine.
o Exine: Hard outer layer. Made up of sporopollenin (highly resistant organic material). It can withstand high temperature and strong acids and alkali. Enzymes cannot degrade sporopollenin.
Exine has apertures called germ pores where sporopollenin is absent.
Pollen grains are preserved as fossils due to the presence of sporopollenin. Exine exhibits patterns and designs.
o Intine: Inner wall. It is a thin and continuous layer made up of cellulose and pectin.
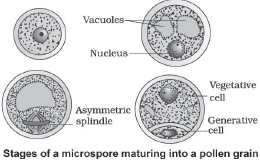
A matured pollen grain contains 2 cells:
o Vegetative cell:
It is bigger, has abundant food reserve and a large irregularly shaped nucleus.
o Generative cell: It is small and floats in the cytoplasm of the vegetative cell. It is spindle shaped with dense cytoplasm and a nucleus.
- Over 60% angiosperms shed their pollen grains at 2-celled stage. In others, generative cell divides mitotically to give 2 male gametes. Thus pollen grains are shed at 3-celled stage.
- The shed pollen grains have to land on the stigma before they lose viability. The viability period of pollen grains is
variable. It depends on temperature and humidity. - Viability of pollen grains of some cereals (rice, wheat etc.) is 30 minutes. Some members of Leguminoseae, Rosaceae
& Solanaceae have viability for months.
Economic importance of pollen grains:
o These are rich in nutrients. Pollen tablets are used as food supplements. Pollen tablets & syrups increase performance of athletes and race horses.
o They are stored for years in liquid nitrogen (-1960 C). They can be used as pollen banks in crop breeding programmes.
o Pollen grains of some plants (e.g. Parthenium or carrot grass) are allergic for some people. It leads to chronic
respiratory disorders (asthma, bronchitis, etc.).
Gynoecium (female reproductive part)
- It may have a single pistil (monocarpellary) or more than one pistil (multicarpellary).
- In multicarpellary, the pistils may be fused together (syncarpous) or free (apocarpous).

Each pistil has three parts:
o Stigma: Landing platform for pollen grains.
o Style: Elongated slender part beneath the stigma.
o Ovary: Basal bulged part. It has ovarian cavity (locule) in which placenta is located. Arising from the placenta are the ovules (megasporangia). Number of ovules in an ovary may be one (wheat, paddy, mango etc.) to many (papaya, water melon, orchids etc.).
Structure of Megasporangium (Ovule):
- Ovule is attached to the placenta by a stalk (funicle).
- Junction between the body of ovule and funicle is called hilum.
- Each ovule has 1 or 2 protective envelopes (integuments) except at the tip where a small opening (micropyle) is present.
- Opposite the micropylar end is the chalaza (basal part).
- Enclosed within the integuments, there is a mass of cells called nucellus. Its cells contain reserve food materials.
- Inside the nucellus is embryo sac (female gametophyte).
- An ovule generally has a single embryo sac formed from a megaspore.
Megasporogenesis:
- It is the formation of megaspores from megaspore mother cell (MMC).
- Ovules generally differentiate a single MMC in micropylar region of the nucellus. It is a large cell containing dense cytoplasm and a prominent nucleus.
- MMC undergoes meiosis to produce 4 megaspores.

Formation of Female gametophyte (embryo sac):
- In majority of flowering plants, one megaspore is functional while the other three degenerates.
- The functional megaspore develops into the female gametophyte. The embryo sac formation from a single
megaspore is called monosporic development. - Nucleus of the functional megaspore divides mitotically to form two nuclei. They move to the opposite poles, forming
2-nucleate embryo sac. - The nuclei again divide two times forming 4-nucleate and 8-nucleate stages of the embryo sac.
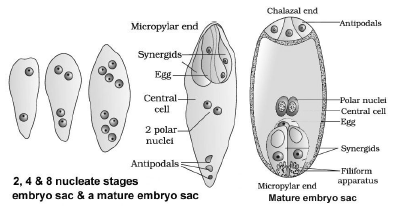
- These divisions are free nuclear, i.e. nuclear divisions are not followed immediately by cell wall formation.
- After the 8-nucleate stage, cell walls are laid down leading to the organization of the typical female gametophyte.
- 6 of the 8 nuclei are surrounded by cell walls and organized into cells. Remaining 2 nuclei (polar nuclei) are situated
below the egg apparatus in the large central cell.
Distribution of cells within the embryo sac:
A typical mature embryo sac is 8-nucleate and 7-celled.
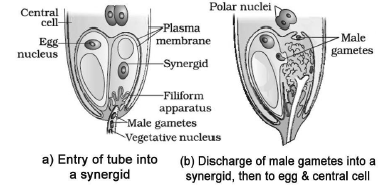
o 3 cells (2 synergids + one egg cell) are grouped at the micropylar end and form egg apparatus.
Synergids have special cellular thickenings at the micropylar tip called filiform apparatus. It helps to guide the pollen tubes into the synergid.
o 3 cells (antipodals) at the chalazal end.
o A large central cell with two polar nuclei.
POLLINATION
It is the transfer of pollen grains from the anther to the stigma of a pistil.
Based on the source of pollen, pollination is 3 types:
a. Autogamy (self-pollination): It is the transfer of pollen grains from the anther to stigma of the same flower.
In flowers with exposed anthers & stigma, complete autogamy is rare. Autogamy in such flowers requires synchrony in pollen release and stigma receptivity. Also, anthers & stigma should be close to each other.
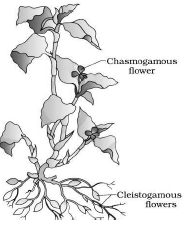
Plants like Viola (common pansy), Oxalis & Commelina produce 2 types of flowers:
• Chasmogamous flowers: They are similar to flowers of other species with exposed anthers and stigma.
• Cleistogamous flowers: They do not open at all.
Anthers & stigma lie close to each other. They are autogamous. When anthers dehisce in the flower buds, pollen grains
come in contact with stigma for pollination.
Cleistogamous flowers produce assured seed-set even in the absence of pollinators.
Cleistogamy leads to inbreeding depression.
b. Geitonogamy: It is the transfer of pollen grains from the anther to the stigma of another flower of the same plant.
It is functionally cross-pollination involving a pollinating agent. But it is genetically similar to autogamy since the pollen grains come from the same plant.
c. Xenogamy: It is the transfer of pollen grains from anther to the stigma of a different plant. It brings genetically different pollen grains to the stigma.
Agents of Pollination
- Abiotic agents (wind & water)
Pollination by wind (anemophily):
- More common abiotic agent.
- Wind pollinated flowers often have a single ovule in each ovary and numerous flowers packed into an inflorescence.
- E.g. Corncob – the tassels are the stigma and style which wave in the wind to trap pollen grains. Wind-pollination is
quite common in grasses. - Ways for effective pollination:
o The flowers produce enormous amount of pollen.
o Pollen grains are light and non-sticky.
o They often possess well-exposed stamens (for easy dispersion of pollens into wind currents).
o Large, feathery stigma to trap air-borne pollen grains.
Pollination by water (hydrophily): - It is quite rare. It is limited to about 30 genera, mostly monocotyledons. E.g. Vallisneria & Hydrilla (fresh water), Zostera (marine sea-grasses) etc.
- But in lower plants, water is a regular mode of transport for the male gametes. Distribution of some bryophytes &
pteridophytes is limited because they need water for the transport of male gametes and fertilisation. - In Vallisneria, the female flower reaches the surface of water by the long stalk and the male flowers or pollen
grains are released on to the surface of water. They are carried by water currents and reach the female flowers. - In sea grasses, female flowers remain submerged in water.
Pollen grains are long and ribbon like. They are carried inside the water and reach the stigma. - The pollen grains of most of the water-pollinated species have a mucilaginous covering to protect from wetting.
- Not all aquatic plants use hydrophily. In most of aquatic plants (water hyacinth, water lily etc.), the flowers emerge
above the level of water for entomophily or anemophily. - Wind and water pollinated flowers are not very colourful and do not produce nectar.
- Biotic agents (animals)
- Majority of flowering plants use animals as pollinating agents. E.g. Bees, butterflies, flies, beetles, wasps, ants,
moths, birds (sunbirds & humming birds) bats, primates (lemurs), arboreal (tree-dwelling) rodents, reptiles (gecko
lizard & garden lizard) etc. - Pollination by insects (Entomophily), particularly bees is more common.
- Often flowers of animal pollinated plants are specifically adapted for a particular species of animal.
- Features of insect-pollinated flowers:
o Large, colourful, fragrant and rich in nectar. Nectar & pollen grains are the floral rewards for pollination.
o Small flowers form inflorescence to make them visible.
o The flowers pollinated by flies and beetles secrete foul odours to attract these animals.
o The pollen grains are generally sticky. - When the animal comes in contact with the anthers and the stigma, its body gets pollen grains. When it comes in contact with the stigma, it results in pollination.
- Some plants provide safe places as floral reward to lay eggs.
E.g. Amorphophallus (It has the tallest flower of 6 feet).
A moth species and the plant Yucca cannot complete their life cycles without each other. The moth deposits its eggs
in the locule of ovary. The flower gets pollinated by moth. The larvae come out of the eggs as seeds start developing. - Many insects consume pollen or nectar without bringing about pollination. They are called pollen/nectar robbers.
Outbreeding Devices
Hermaphrodite flowers can undergo self-pollination.
Continued self-pollination results in inbreeding depression.
To avoid self-pollination (autogamy) and encourage crosspollination, there are some devices in plants:
a. Avoiding synchronization: Here, the pollen is released before the stigma becomes receptive or stigma becomes
receptive before the release of pollen.
b. Arrangement of anther & stigma at different positions.
c. Self-incompatibility: It is a genetic mechanism to prevent self-pollen (from same flower or other flowers of the same plant) from fertilization by inhibiting pollen germination or pollen tube growth in the pistil.
d. Production of unisexual flowers: If male & female flowers are present on the same plant (i.e., monoecious, e.g. castor & maize), it prevents autogamy but not geitonogamy. In dioecious plants (e.g. papaya), male and female flowers are present on different plants (dioecy).
This prevents both autogamy and geitonogamy.
Pollen-pistil Interaction
- It is a process in which pistil recognizes compatible or incompatible pollen through the chemical components produced by them.
- Pistil accepts compatible pollen and promotes postpollination events.
- It rejects incompatible pollen by preventing pollen germination or pollen tube growth.
- Pollen grain germinates on the stigma to produce a pollen tube through one of the germ pores. The contents of pollen grain move into pollen tube.
Pollen tube grows through the tissues of stigma and style and reaches the ovary. - In plants which shed pollen grains at 2-celled condition (a vegetative cell & a generative cell), the generative cell divides into two male gametes during pollen tube growth.
In plants which shed pollen in 3-celled condition, pollen tubes carry 2 male gametes from the beginning. - Pollen tube → ovary → micropyle → ovule → enters one of the synergids through filiform apparatus. Filiform apparatus guides the entry of pollen tube.
- A plant breeder can manipulate pollen-pistil interaction, even in incompatible pollinations, to get desired hybrids.
Artificial hybridisation
It is a crop improvement programme in which desired pollen grains are used for pollination.
Steps:
o Emasculation: Removal of anthers from the bisexual flower bud of female parent before the anther dehisces.
o Bagging: Here, emasculated flowers are covered with a bag (butter paper) to prevent contamination of its stigma
with unwanted pollen.
o Pollination: When stigma attains receptivity, pollen grains collected from male parent are dusted on the stigma.
o Rebagging the flowers. It is allowed to develop the fruits. For unisexual flowers, there is no need for emasculation.
Female flower buds are bagged before the flowers open.
DOUBLE FERTILISATION
- After entering the synergid, the pollen tube releases 2 male gametes into the cytoplasm of the synergid. One male
gamete moves towards the egg cell and fuses with its nucleus (syngamy) to form zygote (diploid). - The other male gamete moves towards the two polar nuclei located in the central cell and fuses with them to produce a triploid primary endosperm nucleus (PEN). As it involves fusion of 3 haploid nuclei, it is called triple fusion.
- Since 2 types of fusions (syngamy & triple fusion) take place in an embryo sac, it is called double fertilisation.
It is an event unique to flowering plants. - The central cell after triple fusion becomes the primary endosperm cell (PEC) and develops into the endosperm while the zygote develops into an embryo.
POST- FERTILISATION: STRUCTURES & EVENTS
Post-fertilisation events: Endosperm & embryo development, maturation of ovule(s) into seed(s) & ovary into fruit.
Endosperm development
- Primary endosperm cell (PEC) divides repeatedly to form a triploid endosperm tissue.
- Endosperm cells are filled with reserve food materials. They are used for nutrition of the developing embryo.
- In common endosperm development, PEN undergoes successive nuclear divisions to give free nuclei (freenuclear
endosperm). Number of free nuclei varies greatly. - Endosperm becomes cellular due to cell wall formation.
- Tender coconut water is a free-nuclear endosperm (made up of thousands of nuclei) and the surrounding white kernel is the cellular endosperm.
Embryo development
- Embryo develops at the micropylar end of the embryo sac where the zygote is situated.
- Most zygotes divide only after the formation of some endosperm. This provides nutrition to developing embryo.
- In monocots & dicots, seeds differ greatly but embryogeny (early embryonic developments) is similar.
- Zygote → Pro-embryo → Globular → Heart-shaped → Mature embryo.

Dicotyledonous embryo
- It has an embryonal axis and 2 cotyledons.
- Portion of embryonal axis above the level of cotyledons is the epicotyl, which terminates with plumule (stem tip).
- The cylindrical portion below the level of cotyledons is hypocotyl that terminates with the radicle (root tip). The
root tip is covered with a root cap.
Monocotyledonous embryo
- They possess only one cotyledon.
- Cotyledon of the grass family is called scutellum.
- It is situated lateral to the embryonal axis. At its lower end, the embryonal axis has the radicle and root cap enclosed in
coleorrhiza (an undifferentiated sheath). - Portion of embryonal axis above the level of attachment of scutellum is the epicotyl. It has a shoot apex and a few leaf
primordia enclosed in coleoptile (a hollow foliar structure).

Seed from Ovule
- Seed is the fertilized ovule formed inside fruits. It is the final product of sexual reproduction.
- It consists of seed coat(s), cotyledon(s) & an embryo axis.
- The cotyledons are simple, generally thick and swollen due to storage food (as in legumes).
- Mature seeds are 2 types:
o Non-albuminous (Ex-albuminous) seeds: Have no residual endosperm as it is completely consumed during
embryo development. E.g. pea, groundnut, beans.
o Albuminous seeds: Retain a part of endosperm. E.g. wheat, maize, barley, castor, coconut. - Occasionally, in some seeds (black pepper, beet etc.) remnants of nucellus are also persistent. It is called perisperm.
- Integuments of ovules harden as tough protective seed coats. It has a small pore (micropyle) through which O2 &
water enter into the seed during germination. - As the seed matures, it becomes dry by reducing water content (10-15 % moisture by mass). The metabolic activity of the embryo slows down. It may enter a state of inactivity (dormancy). Under favourable conditions (moisture, oxygen & suitable temperature), they germinate.

Advantages of seeds:
• Since pollination and fertilisation are independent of water, seed formation is more dependable.
• Better adaptive strategies for dispersal to new habitats. It helps the species to colonize in other areas.
• They have food reserves. So seedlings are nourished until they are capable of photosynthesis.
• The hard seed coat protects the young embryo.
• Being products of sexual reproduction, they generate new genetic combinations and variations.
• Dehydration & dormancy helps to store seeds. It can be used as food throughout year and to raise crop in next season.
Viability of seeds after dispersal:
- In a few species, the seeds lose viability within a few months. Seeds of many species live for several years.
- Some seeds can remain alive for hundreds of years. The oldest is that of a lupine (Lupinus arcticus) excavated from
Arctic Tundra. The seed germinated and flowered after an estimated record of 10,000 years of dormancy. - 2000 years old viable seed is of the date palm (Phoenix dactylifera) discovered during the archeological excavation at King Herod’s palace near the Dead Sea.
Fruit from Ovary
- The ovary develops into a fruit. Transformation of ovules into seeds and ovary into fruit proceeds simultaneously.
- The wall of ovary develops into pericarp (wall of fruit).
- The fruits may be fleshy (e.g. guava, orange, mango, etc.) or dry (e.g. groundnut, mustard etc.).
- Fruits are 2 types:
o True fruits: In this, fruit develops only from the ovary.
Other floral parts degenerate & fall off. E.g. most plants.

APOMIXIS AND POLYEMBRYONY
- Apomixis is the production of seeds without fertilisation.
E.g. Some species of Asteraceae and grasses. - It is a form of asexual reproduction that mimics sexual reproduction.
- In some species, diploid egg cell is formed without reduction division and develops into the embryo without fertilisation.
- In many species (e.g. many Citrus & Mango varieties) some nucellar cells surrounding the embryo sac divide, protrude
into the embryo sac to form embryos. Thus each ovule contains many embryos. Occurrence of more than one embryo in a seed is called polyembryony.
Importance of apomixis in hybrid seed industry - If the seeds collected from hybrids are sown, plants in the progeny will segregate and lose hybrid characters.
- Production of hybrid seeds is costly. So hybrid seeds are also expensive. If the hybrids are made into apomicts, there is no segregation in the hybrid progeny. So farmers can keep on using hybrid seeds to raise new crop.



